© Steven J. Cary, March 2022
About a year ago, I got a bee in my bonnet about eye color on metalmarks in the Las Cruces area. That bee kept after me, leading me to examine every Doña Ana County metalmark photo I could lay my hands on, and some in nearby Texas, too, to see if eye color meant anything. It mattered because Apodemia mejicanus (Sonoran or Mexican Metalmark) and Apodemia duryi (Dury’s Metalmark), the two local varieties, have been a tough pair to completely disentangle when it comes to identification. Butterflyers in that area often reported seeing one or the other, but not always being certain which they saw. I asked Butterflying Cruceños to upload all their relevant photographs to iNaturalist or to BAMONA so I could see as many examples of these metalmarks as possible. Then I promised to get back to them with a full report. My basic conclusions are that, in the study area:
- Ventral wing characters do not help distinguish Apodemia duryi from A. mejicanus.
- Dorsal hindwing basal darkening is helpful in about 85 percent of cases.
- Eye color, green for A. duryi and gray for A. mejicanus, always works, if you can see the eyes.
- Apodemia duryi and A. mejicanus occur on the landscape just as their host plants do.
- Apodemia duryi seems likely to be a distinct species from A. mejicanus in the study area.
Dear Butterfying Cruceños (and any other interested eyes), the full report follows and I welcome any and all comments and suggestions.
Differentiating Two Metalmarks in Doña Ana County, New Mexico
Introduction. Metalmarks (Lepidoptera: Riodinidae) in the genus Apodemia (C. Felder and R. Felder 1865) occur broadly across the American Southwest, where phenotypes proliferate and taxonomy seems to be in slow but regular flux. Given the complexity of this sprawling group, practicality influences most North American butterfly field guide authors to take a general view and to forgo detailed descriptions of the confusing array of local forms and varieties. For example, few modern field guides give clear guidance on how to identify Apodemia duryi (W. H. Edwards 1882) or how to distinguish it from the more widespread, similar and better known Apodemia mejicanus (Behr 1865). Some authorities consider mejicanus and duryi to be independent subspecies of Apodemia mormo (C. Felder and R.. Felder 1859), but in this report they are treated as full species, mostly for convenience. Their actual taxonomic status will come up at the end of this report.
In south-central New Mexico, Doña Ana County is home to Las Cruces, New Mexico’s second largest city, to New Mexico State University, to recently-christened Organ Mountains Desert Peaks National Monument, and to the prominent Organ Mountains, whose jagged, 8990-foot (2741m) summit dominates the horizon for many miles (see Figure 1 below).
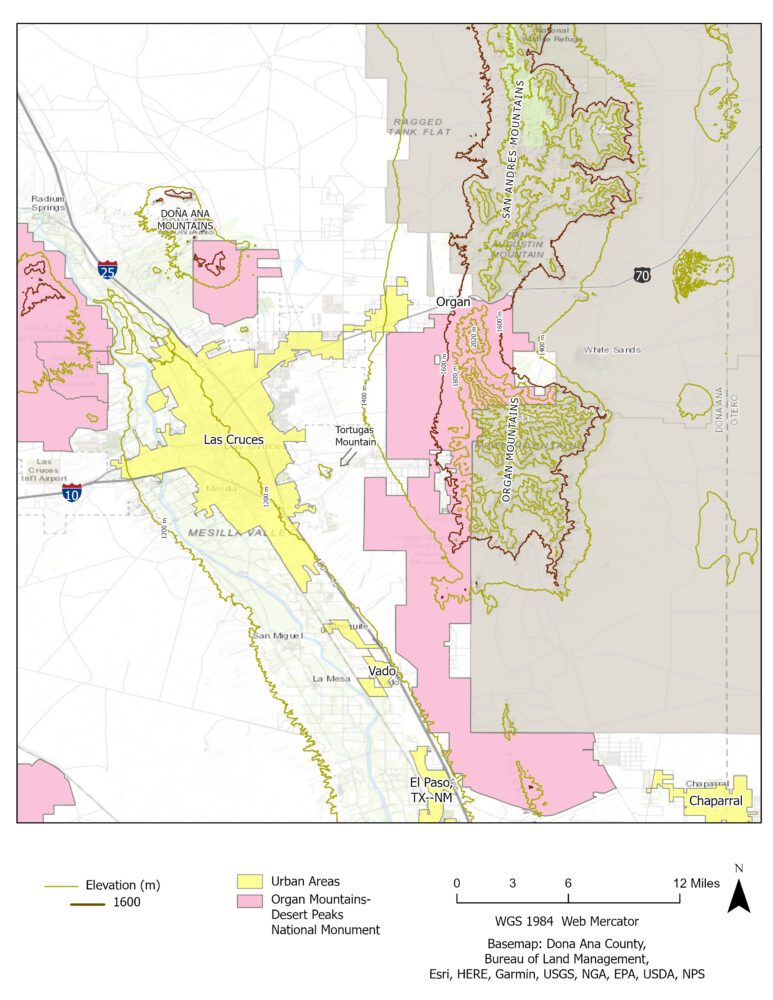
Doña Ana County also is home to four species of metalmarks (see Toliver et al. 2001: 308-316; Cary and Toliver 2021: https://peecnature.org/butterflies-of-new-mexico/metalmarks-riodinidae/). Of the four, Apodemia mejicanus nears the southeastern limit of its expansive western range in the rugged Organ Mountains. Meanwhile, Apodemia duryi is at the northwesternmost known outpost of its Trans-Pecos distribution on the western footslopes of the Organ Mountains. The type locality for A. duryi was stated as “5 miles east of Mesilla” which in my mind translates to Tortugas Mountain, aka “A” Mountain, which is indicated on Figure 1.
Close proximity of these two similar metalmarks on the ground in the study area has placed strain on the means to differentiate them. Most individually observed metalmarks, whether specimens, photos or binocular views, are correctly assigned to either A. mejicanus or A. duryi, but perhaps one in four or five exhibits intermediate characters or traits of both, making it uncertain which species is represented. I and others have had that experience for many years and now, growing legions of amateur naturalists, advances in photographic technology and availability of online citizen science portals have combined to re-focus my attention on this long-smoldering, largely below-the-radar issue concerning proper field identification of A. mejicanus and A. duryi in Doña Ana County.
Tactics traditionally used to differentiate A. duryi from A. mejicanus in the study area have been of inconsistent value, even leading to speculation that the two taxa may be hybridizing. For this inquiry, I drilled down into two popular diagnostic characters related to dorsal and ventral wing colors and patterns. The results, which follow, showed one character to be unfounded and the other to be qualitative and overlapping between the two taxa. Then, I introduce a new diagnostic character, eye color, which seems to be a more effective field mark for distinguishing these two metalmarks.
Methods. To perform analyses and to create maps for this project, I accessed more than 50 photo-illustrated observations posted online as “Approved” at BAMONA (https://www.butterfliesandmoths.org/) or as “Research Grade” at iNaturalist (https://www.inaturalist.org/), all in December 2021. For each individual butterfly depicted (some posts included photos of multiple individuals), I recorded observation date, latitude, longitude, lat/long accuracy when available, plants in association, gender when evident, eye color when evident, and degree of dorsal hindwing (DHW) basal darkening when evident.
The raw geographical data had some issues, as might be expected given that many different observers, recorders and data transmission methods were involved. I found some records entered in more than one website and eliminated duplicates. I found lat/long accuracy to vary from a few meters to thousands of kilometers and I excluded the worst from my analyses. Elevation for each sighting could be obtained by zooming in on the geo-marker overlaid on a terrain map, but it was difficult to know how precisely the original submitter or the transmission software had placed the geo-marker. In the study area’s high-relief terrain, even 100m of lat/long error can produce a grossly inaccurate elevation, so I did not perform analyses with elevation data. I tried not to use data in ways that were unsupported by data quality.
Data related to the butterfly depicted in each image were recorded after close visual examination. Each image brings its unique challenges and constraints. Most images were well-focused, but not all. Most photos showed dorsal or ventral wing surfaces, not both. Some images showed the butterfly at challenging angles. Differential lighting made it impossible to confidently obtain equivalent data from all images. Most photos targeted wing surfaces, often leaving other body parts shaded, not in view, or not in focus. Butterfly gender was plainly discernible in fewer than half the images. Eye color was evident in about 85 percent of images. Degree of DHW basal darkening was evident in about 90 percent of images, reflecting photographers’ general preference for dorsal over ventral views, or the perching and basking preferences of target butterflies. Good ventral views were available in very few images.
Results and Discussion. Our two study subjects (Figure 2) share basic Apodemia features including gray ventral surfaces with white spots and, usually, a thin slice of orange scales. Uppersides share an orange ground color in hues that vary from yellow-orange to burnt orange, with black lines, black submargins, and an overall pattern of white spots that are variably expressed. At a glance, the two metalmarks are the same size and shape, look the same, behave the same and fly the same.
When it comes to differentiating them, traditional tactics for me, and perhaps others, may date back to images and descriptions in William H. Howe’s Butterflies of North America (1975: 270, pl. 47, 77). Dorsally, A. mejicanus was said to have “darker reddish orange markings” whereas A. duryi was said to be “much paler” with “dark markings of the upperside greatly reduced.” Ventrally, A. mejicanus was said to have “some orange” on the hindwing, the amount varying widely across its vast range, whereas A. duryi “lacks orange.” For his descriptions, all Howe could examine was pinned specimens, so it is to be expected that he relied entirely on wing characters when comparing the two species. Subsequent book and field guide authors (e.g., Scott 1986, Opler and Wright 1999, Brock and Kaufman 2003, Glassberg 2017) based their identification guidelines, when offered, on essentially the same wing characters, or they showed an image and let the reader draw her own conclusions.




Figure 2. Photographs of Apodemia mejicanus (left) and Apodemia duryi (right), dorsal (top row) and ventral (bottom row). Cathy Hoyt’s bottom right photo is from Brewster Co., TX. The other three images are from Doña Ana Co., NM.
Howe’s diagnostic instructions were clear, simple and straightforward, so let’s apply them to real world examples. Starting with the ventrum (Figures 2c and 2d), both images show a medium gray ground color splattered with black-edged white spots of various sizes, but in the same basic pattern on each species. A search for orange produces positive results; each image has a small, faint weft of orange woven into scales at the base of the hindwing, in the same place for each. For A. duryi (Figure 2d), this is contrary to Howe’s guidance that the ventrum lacks orange. As it turns out, that individual in Figure 2d is not an oddball. (Photographs of the lectotype of A. duryi can be examined on the Butterflies of America website: http://butterfliesofamerica.com/L/t/Apodemia_duryi_a.htm). Close inspection of the ventrum of the lectotype also reveals the same small, faint weft of orange scaling woven into the pattern at the base of the hindwing.
As regards Apodemia mejicanus (Figure 2c), the original type was lost in the 1906 San Francisco Fire so no one can know if it had orange scaling on the ventrum. Putting aside that key reference point, it certainly seems to be the case that most of what are currently characterized as A. mejicanus colonies in Arizona do have some ventral orange, occasionally quite a bit. Moving east into New Mexico, however, the amount of ventral orange scaling decreases and stabilizes into that single feather visible in Figure 2c. Then one might also ask, will A. mejicanus individuals with zero ventral orange eventually turn up? A recent photograph (Figure 3) of A. mejicanus in the San Andres Mountains, just north of the Organ Mountains, shows a candidate for this honor, but the image has lighting that is not ideal for revealing subtle hindwing features.

To summarize, the traditional diagnostic tool for the ventrum – presence or absence of orange – seems to be poorly founded. Perhaps some A. duryi individuals do in fact lack ventral orange, but the lectotype has some ventral orange and some modern photos of wild individuals also show ventral orange. That ventral orange “feather” is in the same position on A. mejicanus and A. duryi. While some A. duryi may have the orange feather withered to nothing, that also could be true for some A. mejicanus individuals. A more sophisticated analysis of ventral patterns may someday reveal a useful character, but for the time being no ventral character stands out as a reliable means to distinguish A. duryi from A. mejicanus in the study area.
The ventrum “test” was a simple question of presence or absence, but the traditional dorsal test for A. duryi vs. A. mejicanus is less straightforward. Images in Figures 2a and 2b demonstrate that the fundamental arrangement of lines, marks and colors is essentially the same for the two taxa. Each hindwing has a black submargin with a row of small white dots. A larger pattern of white spots overlays dorsal wing surfaces. The hindwing of each has a tangerine orange postmedian region, but basal and median wing regions differ noticeably between the two individuals. For A. duryi (Figure 2b), median and basal areas show an orange pigment that is only faintly smokier than the postmedian tangerine. For A. mejicanus (Figure 2a), the comparable basal region of the dorsal hindwing (DHW) is much darker, almost matte black. Whether that black occurs as a pigment or as a heavy layer of black scales overlaying orange, the difference between these two metalmarks is plain. If all individuals of each taxon were so starkly different in dorsal wing facies, there would be little difficulty differentiating them.
However, examination of multiple individuals across both taxa reveals that the degree of DHW basal darkening varies continuously between completely orange (Figure 2b) and completely black (Figure 2a), which turn out to exemplify opposite ends of a gradational spectrum. A good intermediate example is shown in Figure 4. To which species does one assign individuals like this, with a moderate amount of DHW basal darkening?

This begs the question: is DHW basal darkening an effective and reliable criterion by which to distinguish A. mejicanus from A. duryi? Seeking an answer, I performed a simple analysis. For a total of 32 individuals harvested from online citizen science portals and representing both taxa, undifferentiated, I subjectively scored the intensity of darkening in the DHW basal region as none, slight, moderate, heavy or severe. I used MSExcel to chart the frequency of individuals exhibiting each intensity level, shown as a histogram in Figure 5.
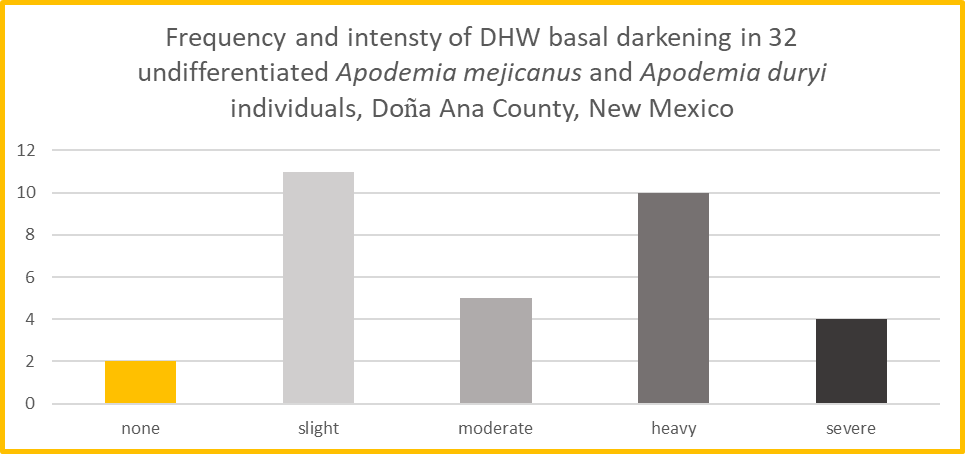
This chart exhibits two distinct frequency peaks. The peak on the left is the cluster of individuals with “slight” darkening, which fits our general perception and expectation for A. duryi. The peak on the right is the cluster of individuals with “heavy” darkening, which matches our general perception and expectation for A. mejicanus. Allowing for some modest variation about each mean, the bimodality of the histogram suggests that DHW basal darkening can be used to distinguish the two metalmarks, at least some of the time.
However, Figure 5 also exhibits a middle bar for “moderate” DHW basal darkening, representing five (15.6%) of our 32 individuals which did not group with either “light” or “heavy.” They are intermediate with regard to DHW basal darkening. This helps to explain the uncertainty some observers experience. Metalmarks with moderate DHW basal darkening are impossible to place with either A. duryi or A. mejicanus based on that character alone.
Collectively then, the dorsal and ventral wing characters traditionally used to distinguish A. duryi from A. mejicanus are not satisfying our identification needs. One is of no use at all and the other only works some of the time. Another character is needed to more effectively differentiate these two butterflies. Recent examination of high-quality color photographs posted online by amateur naturalists shows that the eyes of these two taxa are either gray or green. You can see and compare those hues in the images in Figure 2c and 2d. Try zooming in to get a magnified view.
To test effectiveness of eye color as a potential diagnostic tool to differentiate A. duryi and A. mejicanus in the study area, I identified 28 online observations which showed eye color as well as dorsal wing facies. Then I used MSExcel to augment the chart in Figure 5 and create Figure 6 (below), which distributes eye color along the spectrum of DHW basal darkening, from none to severe.
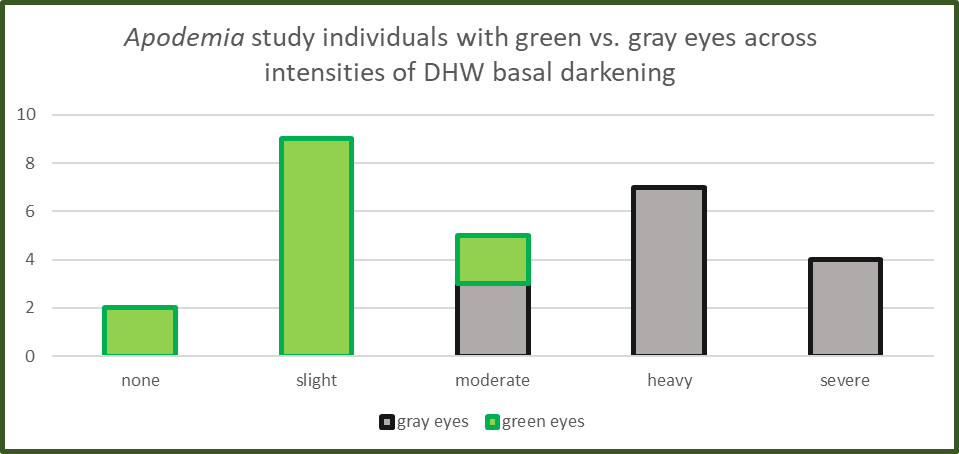
Examination of Figure 6 reveals that it, too, is bimodal, supporting the notion that we are dealing with two distinct taxonomic entities. Green-eyed individuals vary from zero to moderate in terms of DHW basal darkening, with most having a “slight” amount. Gray-eyed individuals cluster about a mean condition of “heavy” DHW basal darkening, while varying from “moderate” to “severe”. A search of the universe of verified online images confirms that A. duryi consistently has green eyes not only in our study area but throughout its entire US range: east to Carlsbad, NM, and south to Big Bend National Park, TX. Similarly, throughout its occurrence in 22 New Mexico counties and regardless of subspecies, A. mejicanus eyes are gray. Armed with that evidence, let’s examine the middle bar in Figure 6, which shows that among individual metalmarks with a moderate or intermediate amount of DHW basal darkening, some have green eyes and some have gray eyes. More directly stated, eye color seems to distinguish A. duryi from A. mejicanus, even for individuals with intermediate DHW basal darkening.
To summarize the identification issues, we can dispense with the ventral orange/no orange test and we now understand that dorsal hindwing basal darkening works most, but not all, of the time. A superior character, eye color, when visible in a photo, in binoculars or to a naked eye, can be used on its own to distinguish green-eyed A. duryi from gray-eyed A. mejicanus in the study area. Notably, green is not an unusual eye color for North American metalmarks. Apodemia palmerii throughout its range and Apodemia mormo in the Four Corners region (at least) also have chartreuse green eyes. The gray eyes of A. mejicanus seem more unique to that species. Eye color should work well in the field if observers and photographers are able to target the head and eyes, in addition to wing surfaces.
Ecology and Biogeography. Over the years, individual Apodemia spp. with intermediate DHW basal darkening have frustrated study area lepidopterists, prompting shoulder shrugs as to identity and fomenting thoughts of hybridization between the two as a potential explanation for the intermediate features. Note, however, that Toliver et al. (2001: 311) asserted unabashedly that “A. duryi does not intergrade with any A. mormo phenotype” by which I presume the authors meant there was no interbreeding between A. duryi and A. mejicanus. They did not explain their reasoning, but perhaps our new clarity about how to tell them apart will help us reason out what those authors intuitively understood.
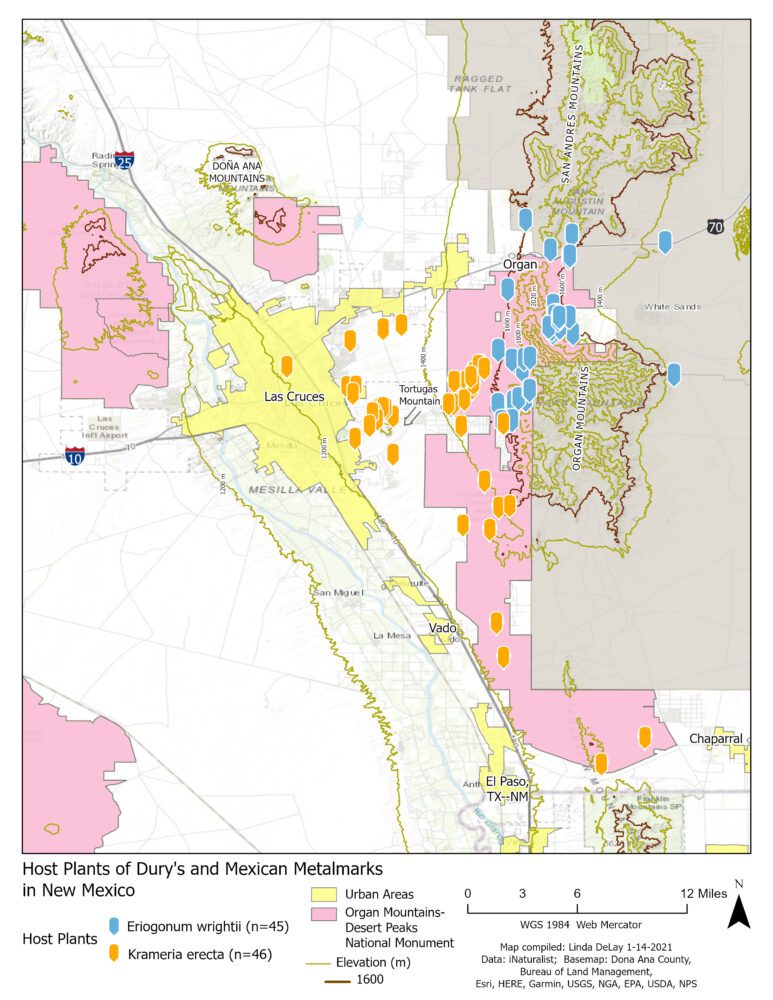
Ecology and biogeography of A. duryi and A. mejicanus in the study area are dictated largely by ecology and biogeography of their respective larval host plants. Almost all metalmark taxa in the A. mormo “superspecies” are hosted exclusively by Eriogonum species. For A. mejicanus, the predominant and perhaps only confirmed larval host throughout southeastern Arizona (Bailowitz and Brock 2022: 119) and southern New Mexico (Cary and Toliver 2022) is the small shrub, Eriogonum wrightii Torr. ex Benth, Polygonaceae (= Wright’s Buckwheat or Bastard Sage). This appears to hold true in the study area, too (see Figure 7).

Notoriously, Apodemia duryi may be the only exception to that widespread Eriogonum “rule.” Throughout its known range, including west Texas, A. duryi is hosted exclusively by a different small shrub, Krameria erecta Willd. ex Schult (= glandulosa = parvifolia; Krameriaceae; Littleleaf Ratany, Range Ratany, Sticky Range Ratany). This host plant association for A. duryi may have been published first by Scott (1986: 348-349), though I suspect it was Gregory S. Forbes, of Las Cruces, who made the initial discovery (see Figure 8 below).

The respective host plants of A. mejicanus and A. duryi occur in different parts of the southern New Mexico landscape. In his accounts for these two common southwestern shrubs, Carter (2012: 228, 364) offered helpful ecological details including the elevations at which they grow. Apodemia duryi’s host, Krameria erecta, is typically found at elevations below 1500m (5000 ft), whereas the A. mejicanus host, Eriogonum wrightii, typically grows above 1500m elevation. Both host plants occur broadly across Doña Ana County, but given their altitudinal preferences, how do they arrange themselves in the study area?
One can visit iNaturalist, search for each plant and see a map for each showing where it has been documented. I wanted to compare distributions of both plants on the same map, so I asked friend and colleague Linda DeLay to take iNaturalist latitude/longitude data and create a map showing both K. erecta and E. wrightii. That map, shown in Figure 9, indicates that these two plants are essentially juxtaposed on the western slope of the Organ Mountains, with little if any overlap. Krameria erecta grows on the lower western footslopes just above the east side of Las Cruces. Eriogonum wrightii occupies slightly higher in the Organ Mountains themselves, uphill to the east. Krameria erecta and E. wrightii exhibit almost zero topographic overlap with each other in the study area. The map in Figure 9 depicts the 1400m and 1600m contours; in English units, the 5000-ft contour lies between the 1400m and 1600m contours.
Ecology of our two metalmarks in the study area also is influenced by their similar adult behaviors. Scott (1986: 349) informs us that each is host-oriented in its various activities. Adults fly within a few feet of the ground surface, bask and perch on the host, and seek mates on or near the host. Adults may nectar at host inflorescences or fly a few dozen meters to seek nectar away from host stands. Neither is known to fly distances to seek mates on hilltops or to patrol for mates along drainages. While some butterfly species are intentionally dispersive and wide-ranging, A. duryi and A. mejicanus exemplify the opposite strategy and rarely wander far from their habitat-specific hosts.
Each metalmark is bound to its caterpillar host plant for purposes of reproduction, so the habitat of the host becomes the habitat of its obligate herbivore metalmark. Under those circumstances, it seems reasonable to expect our metalmarks to exhibit a spatial arrangement in the local landscape similar to that of their hosts. Ms. DeLay, a biologist and GIS specialist, very generously built another map for our use by adding to Figure 9 the known sites for A. mejicanus and A. duryi as documented by BAMONA and iNaturalist. This combined map (Figure 10, below) illustrates that the distribution of each metalmark does indeed mimic the distribution of its larval host. As a result, all confirmed study area reports of gray eyes (= A. mejicanus) are from elevations above 1460m (4800 ft), while all those for green eyes (= A. duryi) are from below 1524m (5000 ft) elevation.
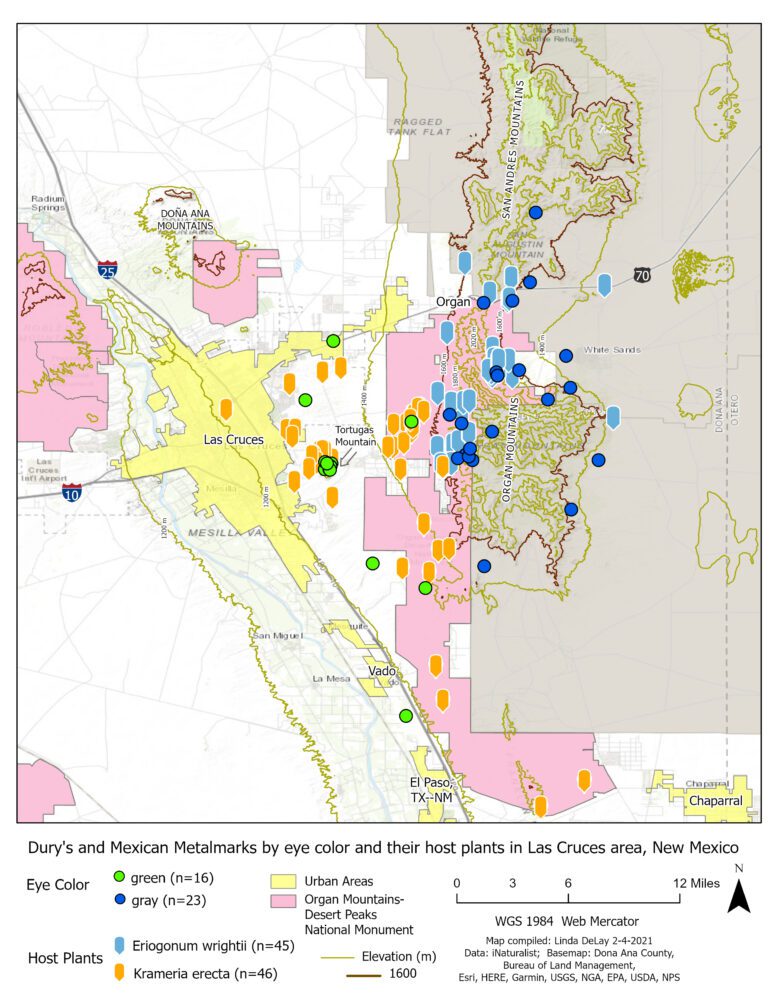
Knowing that distributions of Krameria erecta and Eriogonum wrightii are largely adjacent to each other (Figure 9), and knowing that Apodemia duryi and Apodemia mejicanus retain their distinct identities while mimicking their host plant distributions on the slopes of the Organ Mountains (Figure 10), allows us to reconsider the taxonomic status of the two metalmarks, which for years has been clouded.
Taxonomy. I began this exploration treating A. duryi and A. mejicanus as distinct species, but that was a mere convenience, following the popular Pelham catalog (http://butterfliesofamerica.com/). Multiple opinions are held by various experts and authorities. It may have been Opler and Wright (1999) who first treated Apodemia duryi as a full species in a major book. That apparently was based on recommendations from Gregory S. Forbes and Richard Holland (NABA Names Committee 2001:14), two people who knew that insect as well as anyone at that time. Concurrently, Toliver et al. (2001: 311) also treated A. duryi as a full species, but mejicanus as a subspecies of A. mormo. Field guides published in the recent past (e.g., Brock and Kaufman 2003, Glassberg 2017) treated mejicanus and duryi as independent subspecies with the A. mormo “superspecies” complex. Among current citizen science web platforms, iNaturalist and BAMONA treat them as separate species, while BugGuide (https//bugguide.net) treats duryi as a probable subspecies of A. mejicanus, while calling for more study.
What is the actual relationship between these two taxa? Is one a subspecies of the other? Are they sister subspecies of a particular species? Are they separate and distinct species? The essence of the relationship will come down this: do they successfully breed with each other and produce fully fertile offspring?
While it is currently impossible to demonstrate that duryi and mejicanus do not interbreed, the evidence presented above suggests that Toliver et al. (2001) were correct. Consider the experiment that nature has set up in the study area: Krameria-feeding and Eriogonum-feeding entities butt up against each other with no intervening physical barrier. In such close proximity and with largely concurrent or overlapping flight periods (one bivoltine, the other trivoltine), unrestricted breeding might be expected to produce a well-mixed population that uses both hosts and does not segregate over the landscape by wing facies, eye color, or any other morphological character. That situation should prevail if only one species were involved, regardless of subsidiary subspecies relationships. Instead, the study area displays the opposite: two distinct eye colors, two distinct modes of dorsal wing facies, two distinct larval hosts and two distinct geographic territories. Seems like two species to me.
One could speculate that, located so near each other along an unobstructed slope and with similar behavioral proclivities, occasional pairings of A. duryi with A. mejicanus can be expected, especially where their respective host plants approach or intermingle. Perhaps an occasional metalmark adult would turn up showing intermediate characters including eye color. But the fact that each of our two metalmarks retains its identity while they co-exist side by side is persuasive, if circumstantial, evidence that interspecies pairings are infrequent, that any surviving offspring are not reproductively functional, and that some inherent, organic, species-level boundary exists between duryi and mejicanus in the study area. Results of future genomic analyses within this complicated group would be of great interest.
Admittedly, all of the above deductions, inferences or conclusions are based on a rather contrived data set stapled together from independent observations and subjected to rather simplistic examination. It’s not a rock-solid foundation for sweeping conclusions, so please consider all the above as hypotheses to be investigated further in the coming years. Do green eyes always stay below ~5000 feet, associated with K. erecta, and with “slight” DHW basal darkening? Do gray eyes always stay above ~5000 feet, associate with E. wrightii, and with “heavy” DHW basal darkening? Does this same apparent elevational segregation pertain on south and east slopes of the Organs? In the San Andres or Franklin Mountains? Let’s get details on life histories by rearing eggs of each through to adulthood on their respective host plants. Let’s get more data, more photographs and enrich the BAMONA, BugGuide and iNaturalist holdings for these two butterflies.
One outcome of this inquiry has been a realization that online citizen science platforms and resources can contribute significantly to knowledge of an organism. For centuries, most information about butterflies resided in private and public collections of killed, spread and labeled specimens. All text descriptions or imagery came from those extremely valuable, priceless and essential resources. In recent decades collections of high-quality, digital, color macrophotographs have exploded into millions (billions?) of images. With increasing sophistication of online citizen science portals such as BAMONA, iNaturalist and BugGuide, those photos are gradually transitioning from private into public assets. There will always be a need for specimens to support a wide range of essential biological inquiries including DNA analyses. In this present inquiry, however, it has been satisfying to isolate a potentially important diagnostic trait through examination of photographs accessible on websites. For those who wield cameras in the field instead of nets, it affirms that we can make important contributions, too.
Acknowledgements
Naturalist/photographers Jim Von Loh, Gordon Berman, Rob Wu, Cathy Hoyt, Judy Yellon, Marta Reece, CJ Goin and Elaine Halbedel generously made their images available or otherwise supported this inquiry. I thank Linda DeLay for contributing her considerable map-making skills to this effort. Mike Toliver, Rob Wu and Jim Von Loh reviewed early drafts of this report and offered useful comments and suggestions.
Literature Cited
Bailowitz, Rich, and Jim Brock, 2022. Southeastern Arizona Butterflies. Tucson (AZ): Wheatmark Publ. Co. 356 pp.
Brock, Jim P., and Kenn Kaufman. 2003. Butterflies of North America. Kaufman Focus Guides. New York (NY): Houghton Mifflin Company. 384 pp.
Carter, Jack L. 2012. Trees and Shrubs of New Mexico. Silver City (NM): Mimbres Publishing. 524 pp.
Cary, Steven J., and Michael E. Toliver. 2022. Butterflies of New Mexico, an online photographic guide. https://peecnature.org/butterflies-of-new-mexico/metalmarks-riodinidae/
Glassberg, Jeffrey. 2017. Butterflies of North America. A Swift Guide. 2nd Ed. Princeton (NJ): Princeton University Press. 420 pp.
Howe, William H. 1975. The Butterflies of North America. Garden City (NY): Doubleday & Co. 633 pp.
NABA Names Committee. 2001. Checklist & English Names of North American Butterflies. Second Ed. North American Butterfly Association. 37 pp.
Opler, Paul A., and Amy Bartlett Wright. 1999. Western Butterflies. Peterson Field Guide Series. Houghton Mifflin Co. New York, NY. 540 pp.
Scott, James A. 1986. The Butterflies of North America. Stanford (CA): Stanford University Press. 583 pp.
Toliver, Michael E., Richard Holland, and Steven J. Cary. 2001: Distribution of Butterflies in New Mexico (Lepidoptera: Hesperioidea and Papilionoidea). Third Ed. Albuquerque (NM): R. Holland. 445 pp.
This is very well done! Thanks, Steve!!!
Very insightful! It’s about time somebody unraveled the mysteries of these two taxa in NM- Congratulations! Now, we need to do the same with mejicanus vs. mormo in northern NM-CO…
Thanks Andy, much appreciated. I was just thinking N NM and S CO, too, must be a disease.
mysteries, puzzles, enigmas . . . ain’t life grand?
Thank you, Steve! Very nicely written and so interesting!
thank you Sunnie!!
Hi Steve – the only thing I like better than reading your insightful and disciplined research, analysis, and conclusions for these species this week is the steady, light rain (just now, add some big snowflakes, too) that is currently falling in Las Cruces! This research, so well presented, allows amateurs like me to better understand what attributes to key on while snapping images of butterflies in general and and these metalmarks in particular. Thanks!!
Jim Von Loh
Jim, a steady rain beats me every time. maybe the rain will produce a nice spring bloom of butterflies when it warms up, which should be soon. your discerning eye is very much appreciated. if you see a bloom of Dury’s Metalmarks, please let me know!